
Elektrofizjologia - Electrophysiology
Elektrofizjologia (z greckiego ἥλεκτρον , ēlektron , „bursztyn” [patrz etymologia „elektronu” ]; φύσις , physis , „natura, pochodzenie”; i -λογία , -logia ) jest gałęzią fizjologii, która bada właściwości elektryczne komórki i tkanki. Obejmuje pomiary zmian napięcia lub prądu elektrycznego lub manipulacje na wielu różnych skalach, od białek z pojedynczym kanałem jonowym do całych narządów, takich jak serce . W neuronauce obejmuje pomiary aktywności elektrycznej neuronów , a w szczególności aktywności potencjału czynnościowego. Zapisy wielkoskalowych sygnałów elektrycznych z układu nerwowego , takie jak elektroencefalografia , można również nazwać zapisami elektrofizjologicznymi. Są przydatne do elektrodiagnostyki i monitorowania .

Definicja i zakres
Klasyczne techniki elektrofizjologiczne
Zasada i mechanizmy
Elektrofizjologia to dział fizjologii, który dotyczy szeroko rozumianego przepływu jonów ( prądu jonowego ) w tkankach biologicznych, aw szczególności technik rejestracji elektrycznej, które umożliwiają pomiar tego przepływu. Klasyczne techniki elektrofizjologii polegają na umieszczaniu elektrod w różnych preparatach tkanki biologicznej. Główne typy elektrod to:
- proste przewodniki stałe, takie jak dyski i igły (pojedyncze lub macierzowe, często izolowane poza końcówką),
- ślady na płytkach obwodów drukowanych lub elastycznych polimerach, również izolowane z wyjątkiem końcówki, oraz
- puste rurki wypełnione elektrolitem, takie jak szklane pipety wypełnione roztworem chlorku potasu lub innym roztworem elektrolitu.
Główne przygotowania obejmują:
- organizmy żywe (przykład u owadów ),
- tkanka wycięta (ostra lub hodowana),
- zdysocjowane komórki z wyciętej tkanki (ostre lub hodowane),
- sztucznie wyhodowane komórki lub tkanki, lub
- hybrydy ww.
Elektrofizjologia neuronalna zajmuje się badaniem właściwości elektrycznych komórek i tkanek biologicznych w obrębie układu nerwowego. Dzięki elektrofizjologii neuronalnej lekarze i specjaliści mogą określić, jak zachodzą zaburzenia neuronalne, obserwując aktywność mózgu danej osoby. Aktywność, na przykład, które części mózgu zapalają się w każdej napotkanej sytuacji. Jeśli elektroda ma wystarczająco małą (mikrometry) średnicę, elektrofizjolog może zdecydować się na umieszczenie jej końcówki w pojedynczej celi. Taka konfiguracja umożliwia bezpośrednią obserwację i wewnątrzkomórkową nagrywanie z wewnątrzkomórkowej aktywności elektrycznej z pojedynczej komórki. Jednak ta inwazyjna konfiguracja skraca żywotność komórki i powoduje wyciek substancji przez błonę komórkową. Aktywność wewnątrzkomórkową można również zaobserwować za pomocą specjalnie uformowanej (pustej) szklanej pipety zawierającej elektrolit. W tej technice mikroskopijna końcówka pipety jest dociskana do błony komórkowej, do której ściśle przylega poprzez oddziaływanie szkła i lipidów błony komórkowej. Elektrolit w pipecie może być doprowadzony do ciągłości płynu z cytoplazmą poprzez dostarczenie do pipety impulsu podciśnienia w celu rozerwania małego płata błony otoczonej obrzeżem pipety ( rejestracja całej komórki ). Alternatywnie, ciągłość jonową można ustalić przez „perforację” plastra przez umożliwienie egzogennemu czynnikowi tworzącemu pory w elektrolicie wprowadzenie się do plastra membrany ( zapis perforowanego plastra ). Wreszcie, łatka może pozostać nienaruszona ( nagrywanie łaty ).
Elektrofizjolog może zdecydować o niewstawianiu końcówki do pojedynczej komórki. Zamiast tego końcówkę elektrody można pozostawić w ciągłości z przestrzenią pozakomórkową. Jeśli końcówka jest wystarczająco mała, taka konfiguracja może umożliwiać pośrednią obserwację i rejestrację potencjałów czynnościowych z pojedynczej komórki, co określa się mianem rejestracji pojedynczej jednostki . W zależności od przygotowania i precyzyjnego rozmieszczenia, konfiguracja zewnątrzkomórkowa może wychwycić aktywność kilku pobliskich komórek jednocześnie, co określa się mianem zapisu wielojednostkowego .
Wraz ze wzrostem rozmiaru elektrody zmniejsza się zdolność rozdzielcza. Większe elektrody są wrażliwe tylko na aktywność netto wielu komórek, określaną jako lokalne potencjały pola . Jeszcze większe elektrody, takie jak nieizolowane igły i elektrody powierzchniowe używane przez neurofizjologów klinicznych i chirurgicznych, są wrażliwe tylko na niektóre rodzaje aktywności synchronicznej w populacjach komórek liczących miliony.
Inne klasyczne techniki elektrofizjologiczne obejmują zapis jednokanałowy i amperometrię .
Modalności elektrograficzne według części ciała
Zapis elektrofizjologiczny w ogólności jest czasami nazywany elektrografią (od elektro- + -grafia , „zapis elektryczny”), przy czym tak wytworzony zapis jest elektrogramem. Jednak słowo elektrografia ma inne sensy (m.in. elektrofotografia ), a określone rodzaje zapisu elektrofizjologicznego są zwykle nazywane specyficznymi nazwami, skonstruowanymi na wzór elektro- + [ forma łącząca części ciała ] + -grafia (skrót ExG). W związku z tym słowo elektrogram (nie jest potrzebne dla tych innych zmysłów ) często ma specyficzne znaczenie elektrogramu wewnątrzsercowego, który jest jak elektrokardiogram, ale z kilkoma inwazyjnymi odprowadzeniami (wewnątrz serca), a nie tylko nieinwazyjnymi odprowadzeniami (na skórze). Zapis elektrofizjologiczny do celów diagnostyki klinicznej należy do kategorii badań elektrodiagnostycznych . Różne tryby „ExG” są następujące:
| Modalność | Skrót | Część ciała | Rozpowszechnienie w zastosowaniu klinicznym |
|---|---|---|---|
| elektrokardiografia | EKG lub EKG | serce (w szczególności mięsień sercowy ), ze skórnymi elektrodami (nieinwazyjne) | 1 — bardzo często |
| elektroatriografia | EAG | przedsionkowy mięsień sercowy | 3 — niepospolity |
| elektrowentykulografia | EVG | komorowy mięsień sercowy | 3 — niepospolity |
| elektrogram wewnątrzsercowy | NWZA | serce (w szczególności mięsień sercowy ), z elektrodami wewnątrzsercowymi (inwazyjne) | 2 — dość powszechne |
| elektroencefalografia | EEG | mózg (najczęściej kora mózgowa ) z elektrodami zewnątrzczaszkowymi | 2 — dość powszechne |
| elektrokortykografia | ECoG lub iEEG | mózg (w szczególności kora mózgowa), z elektrodami wewnątrzczaszkowymi | 2 — dość powszechne |
| elektromiografia | EMG | mięśnie całego ciała (zwykle szkieletowe , czasem gładkie ) | 1 — bardzo często |
| elektrookulografia | EOG | oko — cały glob | 2 — dość powszechne |
| elektroretinografia | ERG | oko — konkretnie siatkówka | 2 — dość powszechne |
| elektronystagmografia | ENG | oko —poprzez potencjał rogówkowo-siatkowy | 2 — dość powszechne |
| elektroolfaktografia | EOG | nabłonek węchowy u ssaków | 3 — niepospolity |
| elektroantenografia | EAG | receptory węchowe w antenach stawonogów | 4 — nie dotyczy klinicznie |
| elektrokochleografia | ECOG lub ECochG | ślimak | 2 — dość powszechne |
| elektrogastrografia | JAJKO | mięśnie gładkie żołądka | 2 — dość powszechne |
| elektrogastroenterografia | EGEG | mięśnie gładkie żołądka i jelit | 2 — dość powszechne |
| elektroglottografia | JAJKO | głośnia | 3 — niepospolity |
| elektropalatografia | EPG | podniebienny kontakt języka | 3 — niepospolity |
| elektroarteriografia | EAG | przepływ tętniczy przez potencjał przepływu wykryty przez skórę | 3 — niepospolity |
| elektroblefarografia | EBG | mięsień powiek | 3 — niepospolity |
| elektrodermografia | EDG | skóra | 3 — niepospolity |
| elektrohisterografia | EHG | macica | 3 — niepospolity |
| elektroneuronografia | ENEG lub ENOG | nerwowość | 3 — niepospolity |
| elektropneumografia | EPG | płuca (ruchy klatki piersiowej) | 3 — niepospolity |
| elektrospinografia | ESG | rdzeń kręgowy | 3 — niepospolity |
| elektrowomerografia | EVG | narząd lemieszowo-nosowy | 3 — niepospolity |
Optyczne techniki elektrofizjologiczne
Techniki elektrofizjologiczne optyczne zostały stworzone przez naukowców i inżynierów w celu przezwyciężenia jednego z głównych ograniczeń technik klasycznych. Klasyczne techniki umożliwiają obserwację aktywności elektrycznej w przybliżeniu w jednym punkcie w obrębie objętości tkanki. Techniki klasyczne ujednolicają zjawisko rozproszone. Zainteresowanie przestrzennym rozkładem aktywności bioelektrycznej skłoniło do opracowania cząsteczek zdolnych do emitowania światła w odpowiedzi na ich środowisko elektryczne lub chemiczne. Przykładami są barwniki wrażliwe na napięcie i białka fluorescencyjne.
Po wprowadzeniu jednego lub więcej takich związków do tkanki przez perfuzję, wstrzyknięcie lub ekspresję genu, można obserwować i rejestrować 1- lub 2-wymiarowy rozkład aktywności elektrycznej.
Nagrywanie wewnątrzkomórkowe
Zapis wewnątrzkomórkowy obejmuje pomiar napięcia i/lub prądu w błonie komórki. Aby wykonać zapis wewnątrzkomórkowy, końcówkę cienkiej (ostrej) mikroelektrody należy wprowadzić do komórki, aby można było zmierzyć potencjał błonowy . Zazwyczaj spoczynkowy potencjał błonowy zdrowej komórki będzie wynosił od -60 do -80 mV, a podczas potencjału czynnościowego potencjał błonowy może osiągnąć +40 mV. W 1963 roku Alan Lloyd Hodgkin i Andrew Fielding Huxley zdobyli Nagrodę Nobla w dziedzinie fizjologii lub medycyny za wkład w zrozumienie mechanizmów leżących u podstaw generowania potencjałów czynnościowych w neuronach. Ich eksperymenty obejmowały wewnątrzkomórkowe nagrania z olbrzymiego aksonu kałamarnicy atlantyckiej ( Loligo pealei ) i były jednymi z pierwszych zastosowań techniki "napięcia clamp". Obecnie większość mikroelektrod używanych do rejestracji wewnątrzkomórkowej to mikropipety szklane o średnicy końcówki < 1 mikrometra i rezystancji kilku megaomów. Mikropipety są wypełnione roztworem o podobnym składzie jonowym do płynu wewnątrzkomórkowego komórki. Włożony do pipety drut z chlorowanego srebra łączy elektrycznie elektrolit ze wzmacniaczem i obwodem przetwarzania sygnału. Napięcie mierzone przez elektrodę jest porównywane z napięciem elektrody odniesienia, zwykle srebrnego drutu pokrytego chlorkiem srebra w kontakcie z płynem zewnątrzkomórkowym wokół komórki. Ogólnie rzecz biorąc, im mniejsza końcówka elektrody, tym wyższa jej rezystancja elektryczna , więc elektroda jest kompromisem między rozmiarem (wystarczająco mała, aby przeniknąć pojedynczą komórkę przy minimalnym uszkodzeniu komórki) a rezystancją (na tyle małą, aby małe sygnały neuronowe mogły być z szumów termicznych w końcówce elektrody).
Zacisk napięcia

Technika clampowania napięciowego umożliwia eksperymentatorowi „zaciśnięcie” potencjału komórki na wybranej wartości. Dzięki temu można zmierzyć, ile prądu jonowego przepływa przez błonę komórki przy dowolnym napięciu. Jest to ważne, ponieważ wiele kanałów jonowych w błonie neuronu to kanały jonowe bramkowane napięciem , które otwierają się tylko wtedy, gdy napięcie błony mieści się w określonym zakresie. Pomiary prądu z cęgami napięcia są możliwe dzięki niemal jednoczesnemu cyfrowemu odejmowaniu przejściowych prądów pojemnościowych, które przechodzą, gdy elektroda rejestrująca i błona komórki są ładowane w celu zmiany potencjału komórki.
Cęgi prądowe
Technika current clamp rejestruje potencjał błonowy poprzez wstrzyknięcie prądu do komórki przez elektrodę rejestrującą. W przeciwieństwie do trybu z ograniczeniem napięcia, w którym potencjał błonowy jest utrzymywany na poziomie określonym przez eksperymentatora, w trybie „prądowego zniesienia” potencjał błonowy może się zmieniać, a wzmacniacz rejestruje napięcie generowane przez komórkę samodzielnie lub jako wynik stymulacji. Ta technika służy do badania reakcji komórki na wnikanie do niej prądu elektrycznego; jest to ważne na przykład dla zrozumienia, w jaki sposób neurony reagują na neuroprzekaźniki, które działają poprzez otwieranie błonowych kanałów jonowych .
Większość wzmacniaczy cęgowych zapewnia niewielkie lub żadne wzmocnienie zmian napięcia rejestrowanych z ogniwa. „Wzmacniacz” jest w rzeczywistości elektrometrem , czasami określanym jako „wzmacniacz o wzmocnieniu jedności”; jego głównym celem jest zmniejszenie obciążenia elektrycznego małych sygnałów (w zakresie mV) wytwarzanych przez ogniwa, aby mogły być one dokładnie rejestrowane przez elektronikę o niskiej impedancji . Wzmacniacz zwiększa prąd za sygnałem, jednocześnie zmniejszając rezystancję, przez którą przepływa ten prąd. Rozważmy ten przykład oparty na prawie Ohma: napięcie 10 mV jest generowane przez przepuszczenie 10 nanoamperów prądu przez 1 MΩ rezystancji. Elektrometr zmienia ten „sygnał o wysokiej impedancji” na „sygnał o niskiej impedancji” za pomocą obwodu wtórnika napięcia . Wtórnik napięciowy odczytuje napięcie na wejściu (spowodowane małym prądem przez duży rezystor ). Następnie instruuje obwód równoległy, który ma za sobą duże źródło prądu (sieć elektryczna) i dostosowuje rezystancję tego obwodu równoległego, aby dać to samo napięcie wyjściowe, ale przy niższej rezystancji.
Nagrywanie patch-clamp

Ta technika została opracowana przez Erwina Nehera i Berta Sakmanna, którzy otrzymali Nagrodę Nobla w 1991 roku. Konwencjonalny zapis wewnątrzkomórkowy polega na nakłuciu komórki cienką elektrodą; nagrywanie patch-clamp ma inne podejście. Mikroelektroda typu patch-clamp to mikropipeta o stosunkowo dużej średnicy końcówki. Mikroelektrodę umieszcza się obok komórki i przez mikroelektrodę stosuje się delikatne ssanie w celu wciągnięcia fragmentu błony komórkowej („łaty”) do końcówki mikroelektrody; szklana końcówka tworzy z błoną komórkową „uszczelnienie” o wysokiej odporności. Ta konfiguracja jest trybem „związanym z komórką” i może być wykorzystana do badania aktywności kanałów jonowych obecnych w płatku błony. Jeśli teraz zastosuje się większe ssanie, mały płat membrany w końcówce elektrody może zostać przemieszczony, pozostawiając elektrodę szczelnie przylegającą do reszty ogniwa. Ten tryb „całokomórkowy” umożliwia bardzo stabilny zapis wewnątrzkomórkowy. Wadą (w porównaniu z konwencjonalnym zapisem wewnątrzkomórkowym za pomocą ostrych elektrod) jest to, że płyn wewnątrzkomórkowy komórki miesza się z roztworem wewnątrz elektrody rejestrującej, a zatem niektóre ważne składniki płynu wewnątrzkomórkowego mogą być rozcieńczone. Wariant tej techniki, technika „perforowanej łaty”, stara się zminimalizować te problemy. Zamiast stosowania ssania w celu odsunięcia plastra membrany od końcówki elektrody, możliwe jest również wykonanie małych otworów w plastrze za pomocą środków porotwórczych, aby duże cząsteczki, takie jak białka, mogły pozostać wewnątrz komórki, a jony mogły swobodnie przechodzić przez otwory . Również płat membrany można oderwać od reszty komórki. Takie podejście umożliwia analizę farmakologiczną właściwości membrany plastra.
Ostre nagrywanie elektrody
W sytuacjach, gdy chcemy zarejestrować potencjał wewnątrz błony komórkowej przy minimalnym wpływie na skład jonowy płynu wewnątrzkomórkowego można zastosować ostrą elektrodę. Te mikropipety (elektrody) są znowu podobne do tych do patch clamp wyciągniętych ze szklanych kapilar, ale pory są znacznie mniejsze, więc wymiana jonów między płynem wewnątrzkomórkowym a elektrolitem w pipecie jest bardzo mała. Opór elektryczny elektrody mikropipety zmniejsza się poprzez wypełnienie 2-4M KCl, a nie stężeniem soli, które naśladuje wewnątrzkomórkowe stężenia jonów stosowane w patch clampingu. Często końcówka elektrody jest wypełniona różnego rodzaju barwnikami, takimi jak żółcień Lucyfera, aby wypełnić zarejestrowane komórki w celu późniejszego potwierdzenia ich morfologii pod mikroskopem. Barwniki są wstrzykiwane przez przyłożenie dodatniego lub ujemnego napięcia stałego lub impulsowego do elektrod w zależności od polaryzacji barwnika.
Nagrywanie pozakomórkowe
Nagrywanie pojedynczej jednostki
Elektroda wprowadzona do mózgu żywego zwierzęcia wykryje aktywność elektryczną generowaną przez neurony sąsiadujące z końcówką elektrody. Jeśli elektroda jest mikroelektrodą o wielkości końcówki około 1 mikrometra, elektroda zwykle wykrywa aktywność co najwyżej jednego neuronu. Nagrywanie w ten sposób jest ogólnie nazywane nagrywaniem „pojedynczym”. Zarejestrowane potencjały czynnościowe są bardzo podobne do potencjałów czynnościowych rejestrowanych wewnątrzkomórkowo, ale sygnały są znacznie mniejsze (zwykle około 1 mV). W ten sposób dokonuje się większości zapisów aktywności pojedynczych neuronów u znieczulonych i przytomnych zwierząt. Zapisy pojedynczych neuronów u żywych zwierząt dostarczyły ważnych informacji na temat przetwarzania informacji przez mózg. Na przykład David Hubel i Torsten Wiesel zarejestrowali aktywność pojedynczych neuronów w pierwotnej korze wzrokowej znieczulonego kota i pokazali, jak pojedyncze neurony w tym obszarze reagują na bardzo specyficzne cechy bodźca wzrokowego. Hubel i Wiesel otrzymali Nagrodę Nobla w dziedzinie fizjologii lub medycyny w 1981 roku.
Nagrywanie wielu jednostek
Jeśli końcówka elektrody jest nieco większa, elektroda może rejestrować aktywność generowaną przez kilka neuronów. Ten rodzaj nagrywania jest często nazywany „nagrywaniem wielu jednostek” i jest często używany u przytomnych zwierząt do rejestrowania zmian aktywności w dyskretnym obszarze mózgu podczas normalnej aktywności. Zapisy z jednej lub więcej takich elektrod, które są blisko siebie, można wykorzystać do identyfikacji liczby komórek wokół niej, a także do określenia, które kolce pochodzą z której komórki. Proces ten nazywa się sortowaniem kolców i jest odpowiedni w obszarach, w których istnieją zidentyfikowane typy komórek o dobrze zdefiniowanych charakterystykach kolców. Jeśli końcówka elektrody jest jeszcze większa, na ogół nie można rozróżnić aktywności poszczególnych neuronów, ale elektroda nadal będzie w stanie rejestrować potencjał pola generowany przez aktywność wielu komórek.
Potencjały pola

Zewnątrzkomórkowe potencjały pola są lokalnymi pochłaniaczami lub źródłami prądu, które są generowane przez zbiorową aktywność wielu komórek. Zwykle potencjał pola jest generowany przez jednoczesną aktywację wielu neuronów przez transmisję synaptyczną . Diagram po prawej pokazuje potencjały pola synaptycznego hipokampa. Po prawej stronie dolny ślad pokazuje falę ujemną, która odpowiada spadkowi prądu spowodowanemu dodatnimi ładunkami wchodzącymi do komórki przez postsynaptyczne receptory glutaminianu , podczas gdy górny ślad pokazuje falę dodatnią, która jest generowana przez prąd opuszczający komórkę (na komórce). ciała), aby zakończyć obwód. Aby uzyskać więcej informacji, zobacz potencjał lokalnego pola .
Amperometria
Amperometria wykorzystuje elektrodę węglową do rejestrowania zmian w składzie chemicznym utlenionych składników roztworu biologicznego. Utlenianie i redukcja odbywa się poprzez zmianę napięcia na aktywnej powierzchni elektrody rejestrującej w procesie znanym jako „skanowanie”. Ponieważ niektóre substancje chemiczne w mózgu tracą lub zyskują elektrony przy charakterystycznym napięciu, można zidentyfikować poszczególne gatunki. Amperometria została wykorzystana do badania egzocytozy w układzie nerwowym i hormonalnym. Wiele neuroprzekaźników monoaminowych ; np. noradrenalina (noradrenalina), dopamina i serotonina (5-HT) ulegają utlenieniu. Metodę można również zastosować do komórek, które nie wydzielają utlenialnych neuroprzekaźników poprzez „ładowanie” ich 5-HT lub dopaminą.
Płaski zacisk łatowy
Planar patch clamp to nowatorska metoda opracowana dla wysokowydajnej elektrofizjologii. Zamiast umieszczania pipety na przylegającej komórce, zawiesinę komórek pipetuje się na chip zawierający mikrostrukturyzowany otwór. Pojedyncza komórka jest następnie umieszczana na otworze przez ssanie i powstaje szczelne połączenie (Gigaseal). Geometria planarna oferuje szereg zalet w porównaniu z klasycznym eksperymentem:
- Pozwala na integrację mikroprzepływów , co umożliwia automatyczną aplikację związków do badań przesiewowych kanałów jonowych .
- System jest dostępny dla technik optycznych lub sond skanujących .
- Perfuzji z wewnątrzkomórkowego strony mogą być wykonywane.
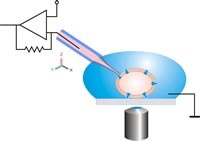
Schematyczny rysunek klasycznej konfiguracji patch clamp. Pipeta typu patch jest przenoszona do celi za pomocą mikromanipulatora pod kontrolą optyczną. Należy unikać ruchów względnych pomiędzy pipetą a komórką, aby utrzymać połączenie komórka-pipeta w stanie nienaruszonym.

Obraz ze skaningowego mikroskopu elektronowego pipety typu patch.
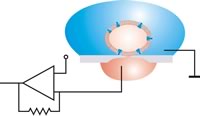
W płaskiej konfiguracji łaty komórka jest pozycjonowana przez odsysanie. Względne ruchy między komórką a otworem można następnie wykluczyć po uszczelnieniu. Stół antywibracyjny nie jest konieczny.

Obraz ze skaningowego mikroskopu elektronowego płaskiego chipa patch clamp. Zarówno pipeta, jak i chip wykonane są ze szkła borokrzemianowego .




Inne metody
Oparte na membranie z podparciem stałym (SSM)
W tym elektrofizjologicznym podejściu proteo liposomy , pęcherzyki błonowe lub fragmenty błony zawierające kanał lub transporter będący przedmiotem zainteresowania są adsorbowane na monowarstwie lipidowej namalowanej na funkcjonalizowanej elektrodzie. Elektroda ta składa się z podłoża szklanego, warstwy chromu, warstwy złota i monowarstwy merkaptanu oktadecylu . Ponieważ pomalowana membrana jest podtrzymywana przez elektrodę, nazywana jest membraną na podłożu stałym. Należy zauważyć, że zaburzenia mechaniczne, które zwykle niszczą biologiczną błonę lipidową, nie wpływają na czas życia SSM. Pojemnościowych elektrod (złożony z SSM i wchłoniętych pęcherzyków) jest tak stabilny mechanicznie, że rozwiązania mogą być szybko wymieniane na jego powierzchni. Ta właściwość pozwala na zastosowanie szybkich skoków stężenia substratu/ligandów w celu zbadania aktywności elektrogenicznej białka będącego przedmiotem zainteresowania, mierzonej poprzez sprzężenie pojemnościowe między pęcherzykami a elektrodą.
Test rozpoznawania bioelektrycznego (BERA)
Test rozpoznawania bioelektrycznego (BERA) to nowatorska metoda oznaczania różnych cząsteczek chemicznych i biologicznych poprzez pomiar zmian potencjału błonowego komórek unieruchomionych w matrycy żelowej. Oprócz zwiększonej stabilności powierzchni styku elektroda-komórka, unieruchomienie pozwala zachować żywotność i funkcje fizjologiczne komórek. BERA jest wykorzystywana głównie w zastosowaniach bioczujników do oznaczania analitów, które mogą oddziaływać z unieruchomionymi komórkami poprzez zmianę potencjału błony komórkowej. W ten sposób, gdy do czujnika dodawana jest próbka dodatnia, następuje charakterystyczna, „sygnaturowa” zmiana potencjału elektrycznego. BERA to podstawowa technologia stojąca za niedawno uruchomionym paneuropejskim projektem FOODSCAN, dotyczącym oceny ryzyka pestycydów i żywności w Europie. BERA jest stosowany do wykrywania ludzkich wirusów (wirusów zapalenia wątroby typu B i C oraz wirusów opryszczki ), weterynaryjnych czynników chorobotwórczych ( wirus pryszczycy , priony i wirus niebieskiego języka ) oraz wirusów roślin (wirusy tytoniu i ogórków) w konkretny, szybki (1–2 minuty), powtarzalny i ekonomiczny sposób. Metoda była również stosowana do wykrywania toksyn środowiskowych, takich jak pestycydy i mykotoksyny w żywności oraz 2,4,6-trichloroanizol w korku i winie, a także do oznaczania bardzo niskich stężeń anionu ponadtlenkowego w próbkach klinicznych .
Czujnik BERA składa się z dwóch części:
- Materiały eksploatacyjne do biorozpoznawania
- Elektroniczne urządzenie odczytujące z wbudowaną sztuczną inteligencją .
Niedawnym postępem jest opracowanie techniki zwanej identyfikacją molekularną poprzez inżynierię membranową (MIME). Technika ta pozwala na budowanie komórek o określonej specyficzności dla praktycznie dowolnej cząsteczki będącej przedmiotem zainteresowania, poprzez osadzenie tysięcy sztucznych receptorów w błonie komórkowej.
Elektrofizjologia obliczeniowa
Chociaż nie stanowią one ściśle pomiaru doświadczalnego, opracowano metody badania właściwości przewodzących białek i biomembran in silico . Są to głównie symulacje dynamiki molekularnej, w których układ modelowy, taki jak dwuwarstwa lipidowa, jest poddawany działaniu napięcia przyłożonego z zewnątrz. Badania z użyciem tych układów umożliwiły zbadanie zjawisk dynamicznych, takich jak elektroporacja błon i translokacja jonów przez kanały.
Zaletą takich metod jest wysoki poziom szczegółowości mechanizmu aktywnego przewodzenia, dzięki z natury wysokiej rozdzielczości i gęstości danych, jakie zapewnia symulacja atomowa. Istnieją poważne wady wynikające z niepewności co do zasadności modelu i kosztu obliczeniowego systemów modelowania, które są wystarczająco duże i w wystarczających skalach czasowych, aby można je było uznać za odtwarzające makroskopowe właściwości samych systemów. Chociaż symulacje atomistyczne mogą mieć dostęp do skal czasowych bliskich lub w zakresie mikrosekund, jest to wciąż kilka rzędów wielkości mniej niż rozdzielczość metod eksperymentalnych, takich jak patch-clamping.
Elektrofizjologia kliniczna
Elektrofizjologia kliniczna to nauka o tym, w jaki sposób zasady i technologie elektrofizjologiczne można zastosować do zdrowia ludzkiego. Na przykład kliniczna elektrofizjologia serca to badanie właściwości elektrycznych, które rządzą rytmem i aktywnością serca. Elektrofizjologia serca może być wykorzystywana do obserwowania i leczenia zaburzeń, takich jak arytmia (nieregularne bicie serca). Na przykład lekarz może wprowadzić do serca cewnik zawierający elektrodę, aby zarejestrować aktywność elektryczną mięśnia sercowego.
Innym przykładem elektrofizjologii klinicznej jest neurofizjologia kliniczna . W tej specjalności medycznej lekarze mierzą właściwości elektryczne mózgu , rdzenia kręgowego i nerwów . Uważa się, że naukowcy tacy jak Duchenne de Boulogne (1806–1875) i Nathaniel A. Buchwald (1924–2006) znacznie rozwinęli dziedzinę neurofizjologii , umożliwiając jej zastosowania kliniczne.
Wytyczne dotyczące raportowania klinicznego
Standardy minimalnych informacji (MI) lub wytyczne dotyczące raportowania określają minimalną ilość metadanych (informacji) oraz danych wymaganych do osiągnięcia określonego celu lub celów w badaniu klinicznym. Rodzina dokumentów z wytycznymi dotyczącymi raportowania „Minimum Information about a Neuroscience research” (MINI) ma na celu zapewnienie spójnego zestawu wytycznych w celu raportowania eksperymentu elektrofizjologicznego. W praktyce moduł MINI zawiera listę kontrolną informacji, które należy podać (np. o zastosowanych protokołach) przy opisywaniu zbioru danych do publikacji.
Zobacz też
- Automatyczny patch clamp
- Bioelektrochemia
- Bioelektromagnetyka
- Elektrofizjologia serca
- Kliniczna elektrofizjologia serca
- Elektrofizjologia kliniczna
- Neurofizjologia kliniczna
- Studium elektrofizjologii
- Równanie Hille'a
- Wieloskalowy format elektrofizjologii
- Neurofizjologia
- Przygotowanie plastra
- Przezskórna elektryczna stymulacja nerwów
Bibliografia
Zewnętrzne linki