
Elektrorecepcja i elektrogeneza -Electroreception and electrogenesis

Elektrorecepcja i elektrogeneza to ściśle powiązane zdolności biologiczne do odbierania bodźców elektrycznych i generowania pól elektrycznych . Oba są używane do lokalizowania ofiary; silniejsze wyładowania elektryczne są używane w kilku grupach ryb do ogłuszania zdobyczy. Zdolności te można znaleźć prawie wyłącznie u zwierząt wodnych lub amfibii, ponieważ woda jest znacznie lepszym przewodnikiemenergii elektrycznej niż powietrza. W pasywnej elektrolokacji obiekty, takie jak zdobycz, są wykrywane poprzez wykrywanie wytwarzanych przez nie pól elektrycznych. W aktywnej elektrolokacji ryby generują słabe pole elektryczne i wyczuwają różne zniekształcenia tego pola wytwarzane przez obiekty przewodzące lub przeciwstawiające się elektryczności. Aktywna elektrolokacja jest praktykowana przez dwie grupy słabo naelektryzowanych ryb , Gymnotiformes (nożowate) i Mormyridae (słoniowate), oraz przez Gymnarchus niloticus , afrykańskiego nożownika. Ryba elektryczna wytwarza pole elektryczne za pomocą narządu elektrycznego , zmodyfikowanego z mięśni ogona. Pole jest nazywane słabym, jeśli wystarcza tylko do wykrycia ofiary, a silnym, jeśli jest wystarczająco silne, aby ogłuszyć lub zabić. Pole może mieć krótkie impulsy, jak u słoniowatych, lub ciągłą falę, jak u scyzoryków. Niektóre silnie elektryczne ryby, takie jak węgorz elektryczny , lokalizują ofiarę, generując słabe pole elektryczne, a następnie silnie rozładowują swoje narządy elektryczne, aby ogłuszyć ofiarę; inne silnie elektryczne ryby, takie jak promień elektryczny , elektrolokują biernie. Obserwatorzy gwiazd są wyjątkowi, ponieważ są silnie elektryczni, ale nie używają elektrolokacji.
Elektroreceptywne ampułki Lorenziniego wyewoluowały na wczesnym etapie historii kręgowców; występują zarówno u ryb chrzęstnoszkieletowych , takich jak rekiny , jak i u ryb kostnoszkieletowych , takich jak coelacanths i jesiotry , i dlatego muszą być starożytne. Większość ryb kostnoszkieletowych wtórnie utraciła ampułki Lorenziniego, ale inne niehomologiczne elektroreceptory wielokrotnie ewoluowały, w tym u dwóch grup ssaków , stekowców ( dziobaka i kolczatki ) oraz waleni ( delfin gujański ).
Historia
.JPG)
W 1678 r. podczas sekcji zwłok rekinów włoski lekarz Stefano Lorenzini odkrył na ich głowach narządy zwane obecnie ampułkami Lorenziniego. Swoje odkrycia opublikował w Osservazioni intorno alle torpedini . Funkcja elektrorecepcyjna tych narządów została ustalona przez RW Murraya w 1960 roku.
W 1921 roku niemiecki anatom Viktor Franz opisał knollenorgany (narządy bulwiaste) w skórze słoniowatych , ponownie bez wiedzy o ich funkcji jako elektroreceptorów.
W 1949 roku ukraińsko-brytyjski zoolog Hans Lissmann zauważył, że afrykański scyzoryk ( Gymnarchus niloticus ) był w stanie pływać do tyłu z taką samą prędkością i z taką samą zręcznością wokół przeszkód, jak wtedy, gdy płynął do przodu, unikając kolizji. W 1950 roku wykazał, że ryba wytwarza zmienne pole elektryczne i że reaguje na każdą zmianę otaczającego ją pola elektrycznego.
Elektrolokacja


Zwierzęta elektroreceptywne używają zmysłu do lokalizowania przedmiotów wokół siebie. Jest to ważne w niszach ekologicznych, gdzie zwierzę nie może polegać na wzroku: na przykład w jaskiniach, w mętnej wodzie iw nocy. Elektrolokacja może być pasywna, wyczuwając pola elektryczne, takie jak te generowane przez ruchy mięśni zakopanej ofiary, lub aktywna, gdy elektrogeniczny drapieżnik wytwarza słabe pole elektryczne, aby umożliwić mu rozróżnienie przewodzących i nieprzewodzących obiektów w jego pobliżu.
Elektrolokacja pasywna
W pasywnej elektrolokacji zwierzę wyczuwa słabe pola bioelektryczne generowane przez inne zwierzęta i wykorzystuje je do ich lokalizacji. Te pola elektryczne są generowane przez wszystkie zwierzęta z powodu aktywności ich nerwów i mięśni. Drugim źródłem pól elektrycznych u ryb jest pompa jonowa związana z osmoregulacją błony skrzelowej . Pole to jest modulowane przez otwieranie i zamykanie pyska i szczelin skrzelowych. Bierna elektrorecepcja zwykle opiera się na receptorach ampułkowych, takich jak ampułki Lorenziniego, które są wrażliwe na bodźce o niskiej częstotliwości, poniżej 50 Hz. Receptory te mają wypełniony galaretką kanał prowadzący od receptorów czuciowych do powierzchni skóry.
Aktywna elektrolokacja


W aktywnej elektrolokacji zwierzę wyczuwa otaczające środowisko, generując słabe pola elektryczne (elektrogeneza) i wykrywając zakłócenia w tych polach za pomocą narządów elektroreceptorów. To pole elektryczne jest generowane przez wyspecjalizowany narząd elektryczny składający się ze zmodyfikowanych mięśni lub nerwów. Zwierzęta, które używają aktywnego elektrorecepcji, obejmują słabo elektryczne ryby , które albo generują małe impulsy elektryczne (określane jako „typ impulsowy”), jak u Mormyridae, albo wytwarzają quasi- sinusoidalne wyładowanie z narządu elektrycznego (określane jako „typ falowy”). , jak u Gymnotidae.
Wiele z tych ryb, takich jak Gymnarchus i Apteronotus , utrzymuje swoje ciało raczej sztywne, pływając do przodu lub do tyłu z równą łatwością dzięki falistym płetwom , które rozciągają się przez większość długości ich ciała. Pływanie do tyłu może pomóc im w wyszukiwaniu i ocenie ofiary za pomocą wskazówek elektrosensorycznych. Eksperymenty przeprowadzone przez Lannoo i Lannoo w 1993 roku potwierdzają propozycję Lissmanna, że ten styl pływania z prostymi plecami działa skutecznie, biorąc pod uwagę ograniczenia aktywnej elektrolokacji. Apteronotus może wybierać i łapać większe rozwielitki spośród mniejszych i nie dyskryminuje sztucznie zaciemnionych pcheł wodnych, w obu przypadkach ze światłem lub bez.
Ryby te wytwarzają potencjał zwykle mniejszy niż jeden wolt (1 V). Ryby słabo elektryczne mogą rozróżniać obiekty o różnych wartościach rezystancji i pojemności , co może pomóc w identyfikacji obiektów. Aktywny elektrorecepcja zwykle ma zasięg około jednej długości ciała, chociaż obiekty o impedancji elektrycznej podobnej do impedancji otaczającej wody są prawie niewykrywalne.
Aktywna elektrolokacja opiera się na bulwiastych elektroreceptorach, które są wrażliwe na bodźce o wysokiej częstotliwości (20-20 000 Hz ). Receptory te mają luźną zatyczkę komórek nabłonkowych , która pojemnościowo łączy komórki receptorów czuciowych ze środowiskiem zewnętrznym. Słonie (Mormyridae) z Afryki mają w skórze bulwiaste elektroreceptory znane jako Knollenorgans i Mormyromasts.
Słonie emitują krótkie impulsy, aby zlokalizować swoją ofiarę. Obiekty pojemnościowe i rezystancyjne inaczej wpływają na pole elektryczne, umożliwiając rybom lokalizowanie obiektów różnego typu w odległości mniej więcej długości ciała. Obiekty rezystancyjne zwiększają amplitudę impulsu; obiekty pojemnościowe wprowadzają zniekształcenia.
![Elektrolokacja obiektów pojemnościowych i rezystancyjnych u słoni. Ryba emituje krótkie impulsy z narządu elektrycznego; jego elektroreceptory wykrywają sygnały zmodyfikowane przez właściwości elektryczne otaczających go obiektów.[1]](https://upload.wikimedia.org/wikipedia/commons/thumb/f/f2/Electroreception_of_Capacitative_and_Resistive_Objects_in_Elephantfish.svg/447px-Electroreception_of_Capacitative_and_Resistive_Objects_in_Elephantfish.svg.png)
Elektrolokacja obiektów pojemnościowych i rezystancyjnych u słoni. Ryba emituje krótkie impulsy z narządu elektrycznego; jego elektroreceptory wykrywają sygnały zmodyfikowane przez właściwości elektryczne otaczających go obiektów.
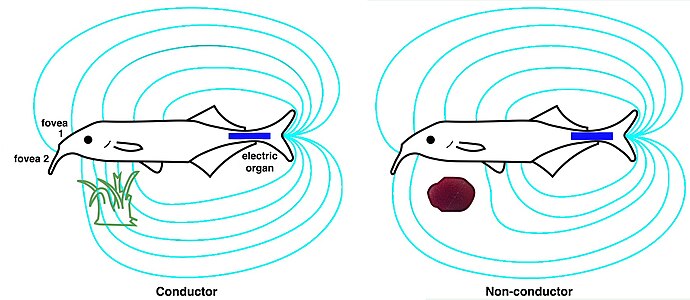
W przypadku słoniowatych narząd elektryczny w ogonie (niebieski) generuje pole elektryczne (cyjan). Jest to wykrywane przez elektroreceptory w skórze, w tym dwa dołki elektryczne (dołki) do aktywnego wyszukiwania i sprawdzania obiektów. Pokazane są zniekształcenia pola powodowane przez dwa różne rodzaje obiektów: roślinę, która przewodzi lepiej niż woda (zielona) i nieprzewodzący kamień (brązowy).
![Elektrolokacja obiektów pojemnościowych i rezystancyjnych u słoni. Ryba emituje krótkie impulsy z narządu elektrycznego; jego elektroreceptory wykrywają sygnały zmodyfikowane przez właściwości elektryczne otaczających go obiektów.[1]](https://wikipedia.org/wiki/File:Electroreception_of_Capacitative_and_Resistive_Objects_in_Elephantfish.svg)

Gymnotiformes , w tym szklany scyzoryk ( Sternopygidae ) i węgorz elektryczny ( Gymnotidae ), różnią się od Mormyridae emitowaniem ciągłej fali, zbliżonej do fali sinusoidalnej, z ich organów elektrycznych. Podobnie jak u Mormyridae, generowane pole elektryczne umożliwia im dokładne rozróżnianie obiektów pojemnościowych i rezystancyjnych.

Elektrolokacja obiektów pojemnościowych i rezystancyjnych w szklanych scyzorykach. |

Narządy elektryczne węgorza elektrycznego zajmują większą część jego ciała. |
Elektrokomunikacja
.jpg)
Słabo elektryczne ryby mogą komunikować się poprzez modulację generowanego przez nie kształtu fali elektrycznej . Mogą to wykorzystać do przyciągnięcia partnerów i pokazów terytorialnych. Sumy elektryczne często wykorzystują swoje wyładowania elektryczne do odstraszania innych gatunków ze swoich schronień, podczas gdy w przypadku własnego gatunku zrytualizowane walki z pokazami otwartych ust i czasami ukąszeniami, ale rzadko używają wyładowań organów elektrycznych.
Kiedy dwa szklane scyzoryki (Sternopygidae) zbliżają się do siebie, oba osobniki zmieniają częstotliwość wyładowań w odpowiedzi unikania zagłuszania .
W tęponosych nożowcach, Brachyhypopomus , wzór wyładowania elektrycznego jest podobny do niskonapięciowego wyładowania elektrolokacyjnego węgorza elektrycznego , Electrophorus . Przypuszcza się, że jest to batesowska mimikra silnie chronionego węgorza elektrycznego. Samce Brachyhypopomus wytwarzają ciągły elektryczny „buczenie”, aby przyciągnąć samice; pochłania to 11–22% ich całkowitego budżetu energetycznego, podczas gdy elektrokomunikacja kobiet zużywa tylko 3%. Duże samce wytwarzały sygnały o większej amplitudzie, które są preferowane przez samice. Koszt dla samców jest zmniejszony przez rytm dobowy , przy czym większa aktywność zbiega się z nocnymi zalotami i tarłem, a mniejsza w innym czasie.
Ryby polujące na ryby elektrolokujące mogą „podsłuchiwać” wyładowania ofiary, aby je wykryć. Elektroreceptywny sum afrykański ostrozębny ( Clarias gariepinus ) może w ten sposób polować na słabo elektrycznego mormyrida Marcusenius macrolepidotus . To spowodowało, że ofiara w ewolucyjnym wyścigu zbrojeń rozwinęła bardziej złożone sygnały o wyższej częstotliwości, które są trudniejsze do wykrycia.
Niektóre zarodki i młode rekinów „zamarzają”, gdy wykryją charakterystyczny sygnał elektryczny swoich drapieżników.
Ewolucja i rozmieszczenie taksonomiczne
U kręgowców pasywna elektrorecepcja jest cechą przodków , co oznacza, że była obecna u ich ostatniego wspólnego przodka. Mechanizm przodków nazywa się elektrorecepcją ampułkową, od nazwy zaangażowanych narządów receptywnych, ampułki Lorenziniego . Wyewoluowały one z mechanicznych czujników linii bocznej i występują w rybach chrzęstnoszkieletowych ( rekinach , płaszczkach i chimerach ) , dwudysznych , bichirach , celakantach , jesiotrach , wiosłonosach , salamandrach wodnych i jesiennych . Wydaje się, że ampułki Lorenziniego zostały utracone na wczesnym etapie ewolucji ryb kostnoszkieletowych i czworonogów , chociaż dowody na brak w wielu grupach są niekompletne i niezadowalające. Tam, gdzie w tych grupach występuje elektrorecepcja, została ona wtórnie nabyta w trakcie ewolucji, przy użyciu narządów innych niż i niehomologicznych z ampułkami Lorenziniego.
Narządy elektryczne ewoluowały co najmniej osiem razy, z których każdy tworzył klad : dwukrotnie podczas ewolucji ryb chrzęstnoszkieletowych, tworząc elektryczne łyżwy i płaszczki, i sześć razy podczas ewolucji ryb kostnoszkieletowych. Grupy pasywnie elektrolokujące, w tym te, które poruszają głowami, aby kierować swoimi elektroreceptorami, są pokazane bez symboli. Gatunki nieelektrolokujące nie są pokazane. Ryby aktywnie elektrolokujące są oznaczone małą żółtą błyskawicą ![]() i charakterystycznymi przebiegami wyładowań. Ryby zdolne do rażenia prądem są oznaczone czerwoną błyskawicą
i charakterystycznymi przebiegami wyładowań. Ryby zdolne do rażenia prądem są oznaczone czerwoną błyskawicą ![]() .
.
| kręgowce |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

.jpg)

Ryby chrzęstne
Rekiny i płaszczki ( Elasmobranchii ) polegają na elektrolokacji przy użyciu ampułek Lorenziniego w końcowych fazach swoich ataków, o czym świadczy silna reakcja żerowania wywołana przez pola elektryczne podobne do tych u ich ofiar. Rekiny są najbardziej wrażliwymi elektrycznie znanymi zwierzętami, reagującymi na pola prądu stałego o natężeniu zaledwie 5 nV/cm.
Oścista ryba
Dwie grupy teleostów są słabo elektryczne i aktywnie elektroreceptywne: neotropikalne scyzoryki ( Gymnotiformes ) i afrykańskie słonie ( Notopteroidei ), umożliwiające im nawigację i znajdowanie pożywienia w mętnej wodzie. Gymnotiformes obejmuje węgorza elektrycznego , który oprócz stosowania przez grupę niskonapięciowej elektrolokacji, jest w stanie generować wstrząsy elektryczne o wysokim napięciu, aby ogłuszyć swoją ofiarę. Tak potężna elektrogeneza wykorzystuje duże narządy elektryczne zmodyfikowane z mięśni. Składają się one ze stosu elektrocytów, z których każdy może generować małe napięcie; napięcia są skutecznie sumowane ( szeregowo ), aby zapewnić potężne wyładowanie organów elektrycznych.
stekowce

Monotremy , w tym dziobak półwodny i kolczatki lądowe, są jedyną grupą ssaków, które rozwinęły elektrorecepcję. Podczas gdy elektroreceptory u ryb i płazów wyewoluowały z mechanosensorycznych narządów linii bocznej, te u stekowców są oparte na gruczołach skórnych unerwionych przez nerwy trójdzielne. Elektroreceptory stekowców składają się z wolnych zakończeń nerwowych zlokalizowanych w gruczołach śluzowych pyska . Spośród stekowców dziobak ( Ornithorhynchus anatinus ) ma najostrzejszy zmysł elektryczny. Dziobak lokalizuje swoją ofiarę za pomocą prawie 40 000 elektroreceptorów ułożonych wzdłuż dzioba w paski od przodu do tyłu. Układ jest wysoce kierunkowy, najbardziej wrażliwy na boki i poniżej. Wykonując krótkie, szybkie ruchy głową zwane sakkadami , dziobaki dokładnie lokalizują swoją ofiarę. Wydaje się, że dziobak wykorzystuje elektrorecepcję wraz z czujnikami ciśnienia do określenia odległości do ofiary na podstawie opóźnienia między pojawieniem się sygnałów elektrycznych a zmianami ciśnienia w wodzie.
Zdolności elektrorecepcyjne czterech gatunków kolczatek są znacznie prostsze. Kolczatki długodziobe (rodzaj Zaglossus ) mają około 2000 receptorów, podczas gdy kolczatki krótkodziobe ( Tachyglossus aculeatus ) mają ich około 400, blisko końca pyska. Różnicę tę można przypisać ich siedlisku i metodom karmienia. Zachodnie kolczatki długodziobe żywią się dżdżownicami w ściółce liściastej w lasach tropikalnych, wystarczająco wilgotnej, aby dobrze przewodzić sygnały elektryczne. Kolczatki krótkodziobe żywią się głównie termitami i mrówkami , które żyją w gniazdach na obszarach suchych; wnętrza gniazd są przypuszczalnie wystarczająco wilgotne, aby zadziałał elektrorecepcja. Eksperymenty wykazały, że kolczatki można nauczyć reagowania na słabe pola elektryczne w wodzie i wilgotnej glebie. Przypuszcza się, że zmysł elektryczny kolczatki jest ewolucyjną pozostałością po przodku podobnym do dziobaka.
Delfiny
Delfiny wyewoluowały elektrorecepcję w strukturach innych niż ryby, płazy i stekowce . Bezwłose krypty wibrysowe na mównicy delfina gujańskiego ( Sotalia guianensis ), pierwotnie związane z wąsami ssaków, są zdolne do elektrorecepcji już od 4,8 μV / cm, wystarczającej do wykrycia małych ryb. Jest to porównywalne z czułością elektroreceptorów dziobaka.
Pszczoły
Do niedawna elektrorecepcja była znana tylko u kręgowców . Ostatnie badania wykazały, że pszczoły potrafią wykryć obecność i wzór ładunku elektrostatycznego na kwiatach.
Zobacz też
Bibliografia
Dalsza lektura
- Bullock, Theodore Holmes (2005). Elektrorecepcja . Nowy Jork: Springer . ISBN 978-0-387-23192-1. OCLC 77005918 .